RNA干渉経路
tetsu
遺伝子発現制御理論物理学研究室

名古屋大学に着任した当時は複合材料を研究する部署にいたため、遺伝子発現制御の研究をする時間は基本的には土日しかとれませんでした。
しかし、朝6~7時くらいに大学に来て、NCCの会議が始まる前に論文を読んでいました。
そのほとんどは、後ほど説明するHi-Cという方法を用いて、DNAの構造を測定する方法でした。
何か新しい発見につながりそうなことが書いてあるという直感はあるのですが、Hi-Cが何を測っているのかいまいちわからず、またその結果が何を表しているのかということもよくわからず、悶々とした日々を過ごしていました。
当時は知り合いの分子生物学者などいなかったので、聞くこともできませんでしたしね。
同時並行でクロマチンブラシの相分離の研究をしていました。
ある時、何かの用事でSchiesselさんとメールを交換したのですが、ループ押し出しという面白い現象があって、その研究をしてみたいと言っていました。
なんだそれ?ということで調べ始めたのが、ループ押し出しの研究を行うきっかけでした。
真核生物のDNAは、ヒストンというタンパク質と複合体(クロマチン)を形成します。
電子顕微鏡でクロマチンを見ると、数珠状の構造をしていて、その珠1つをヌクレオソームと言います。
ヌクレオソームは、ヒストン8つ(8量体)の周りにDNAが巻き付いた構造を取っています。
真核生物のDNAは非常に長い場合が多いですが、1本分のスケールのクロマチンを染色体と呼びます。
新しい細胞は、細胞分裂によって生まれ、成長し、また分裂していきます。
細胞分裂が起こる期間を分裂期、分裂期が終わってから次の分裂期が始まるまでの期間を間期と言います。
間期+分裂期1回分の期間(つまり、新しい細胞ができてから分裂するまでの期間)を細胞周期と言います。
細胞周期は、ヒトで24時間程度、マウスで12時間程度です。
間期と分裂期では、染色体の構造がずいぶん変わります。
分裂期における染色体の構造は、別の記事で解説します。
ヒトなどの高等生物の場合、間期には、それぞれの染色体が核内の決まった位置を占める染色体テリトリー(chromosome territories)という構造を形成します[1]。
英語で良ければ、染色体テリトリーの発見にDNA FISHという技術が役に立ったことに関する一般向けの記事があります[2]。
物理学では、染色体テリトリーは、クロマチンの絡み合いによって形成されると信じられてきました[3,4]。
しかし、現在では、トポイソメラーゼIIというクロマチンの絡み合いをほどく活性を持つ酵素があることが分かっていますし、染色体テリトリーの形成はコンデンシンIIというタンパク質によることも分かっています[5]。
物理学の素朴なアイデアは、なかなか生物系では通用しないんですよね。
このような事例を見ると、少なくとも現時点での分野の知識では、実験分子生物学者との共同研究をして注意深く実験を見ないと難しそうです。
最大のスケールの構造である染色体テリトリーと、最小スケールのヌクレオソームとの間のスケールでのクロマチンの構造は、長らく謎のままでした。
その活路を見出したのが、次にご紹介するHi-C法です。
ヌクレオソームも染色体テリトリーも、顕微鏡で見ることにより発見された構造です。
Hi-C法はそれとは非常に異なるアプローチで染色体の構造を探ります[6]。
この方法では、まず、染色体を化学薬品で急速に架橋します。
架橋に使う分子は低分子ですので、もともと近くにあった2つのDNA部位が架橋されることになります。
どのDNA領域が架橋されたかということは、シーケンサーを使って塩基対を読むことで特定することができます。
これを多数の同種の細胞に対して行います。
この操作を行った細胞数の中で、特定の2つのDNA部位が架橋されていたものの割合が、その2つの部位が近接する頻度(接触頻度、または、contact frequency)となります。
Hi-C法では、すべてのDNA部位のペアについて接触頻度を測ることができます。
接触頻度から染色体の構造を推定することができます。
ペアを考えるDNA部位の長さを小さくするほど解像度が上がります。
そのためには、架橋剤を工夫する必要があるようです。
その後、解像度を上げたMicro-C[7,8]、接触頻度を1細胞で測れるようにしたSingle-Cell Hi-C[9]、切断したDNAを丸ごと液滴に封じ込め、2つ以上のDNA部位の接触まで測れるようにしたChIA-Drop[10]などの改良版、拡張版も開発されました。
Hi-Cを使って最初に分かったことは、1Mbps(塩基対100万個分の長さ)程度の解像度で見ると、染色体は二種類の領域のモザイク構造(つまり、二種類の領域が交互につながった構造)と見ることができるということです[6]。
その二種類の領域をA領域とB領域と呼ぶことにすると、A領域とA領域、および、B領域とB領域の接触頻度は比較的高いのですが、A領域とB領域の間の接触頻度が低くなっています。
つまり、A領域とB領域は、別々の区画(コンパートメント)を作っているということです。
まんまですが、それぞれを、AコンパートメントとBコンパートメントと呼びます。
論文[6]で仮に置かれているA領域には転写活性に関わるヒストンテイル修飾が観察され、B領域には転写抑制に関わるヒストンテイル修飾が観察されています。
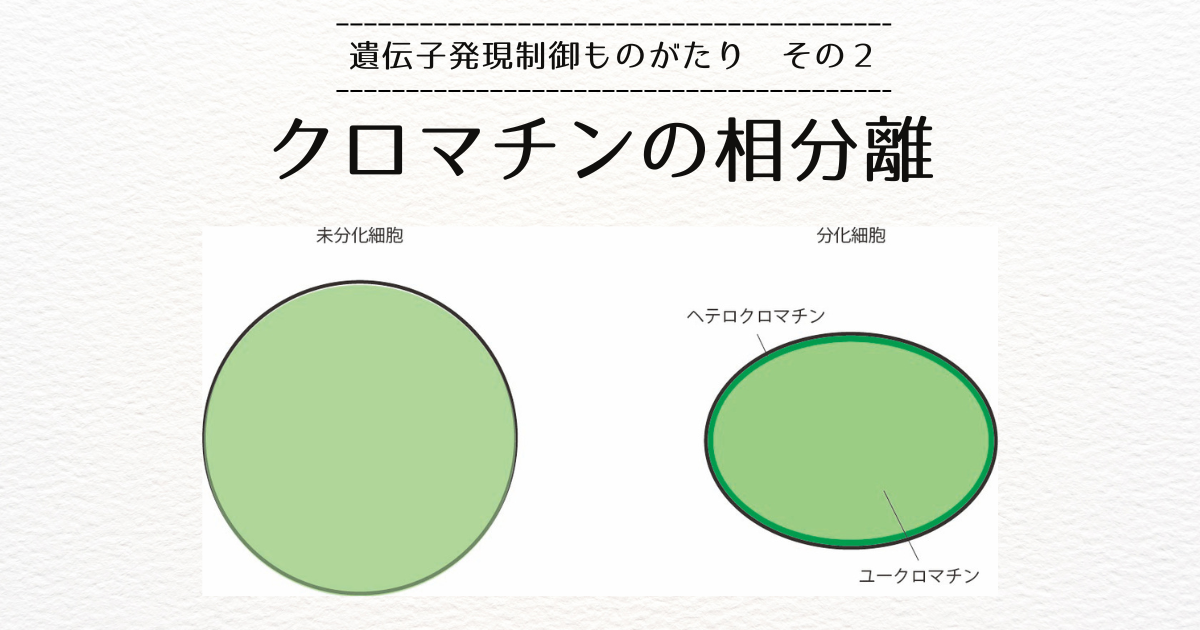
クロマチンの相分離と状況は非常に似通ってます。
クロマチンの相分離では、ユークロマチン(Aコンパートメントに対応)は核の中央に配置し、ヘテロクロマチン(Bコンパートメントに対応)は核膜付近に局在化するということを言いました。
Hi-Cで測られているコンパートメントは、ずっと小さいはずなので、もっと細かく見てもユークロマチンとヘテロクロマチンが分離していることを示唆しています。
Hi-C実験の結果は、コンパートメント以外にもクロマチンの構造について重要な示唆を与えます。
クロマチンは高分子ですので、隣接する部位の接触頻度は高くなっています。
1 kbps(塩基対1000個分の長さ) – 1Mbps(塩基対100万個分の長さ)くらいに分解能を高くしてみると、接触頻度が高い部分がブロック化していることが分かります[11,12]。
どういうことかというと、領域内部のクロマチン部位間の接触頻度が高くなっていますが、その領域内の部位と領域外の部位は接触頻度が高いような領域(ドメイン)に分けられているわけです。
この領域をトポロジカルドメイン(Topologically Associated Domains、略してTAD)と呼びます。
トポロジカルドメイン内部の接触頻度を見てみますと、ドメインの両端間の接触頻度が高くなっていることが分かります[13]。
この結果から、トポロジカルドメインはクロマチンのループ、あるいは、それが集まったものであることを示唆しています。
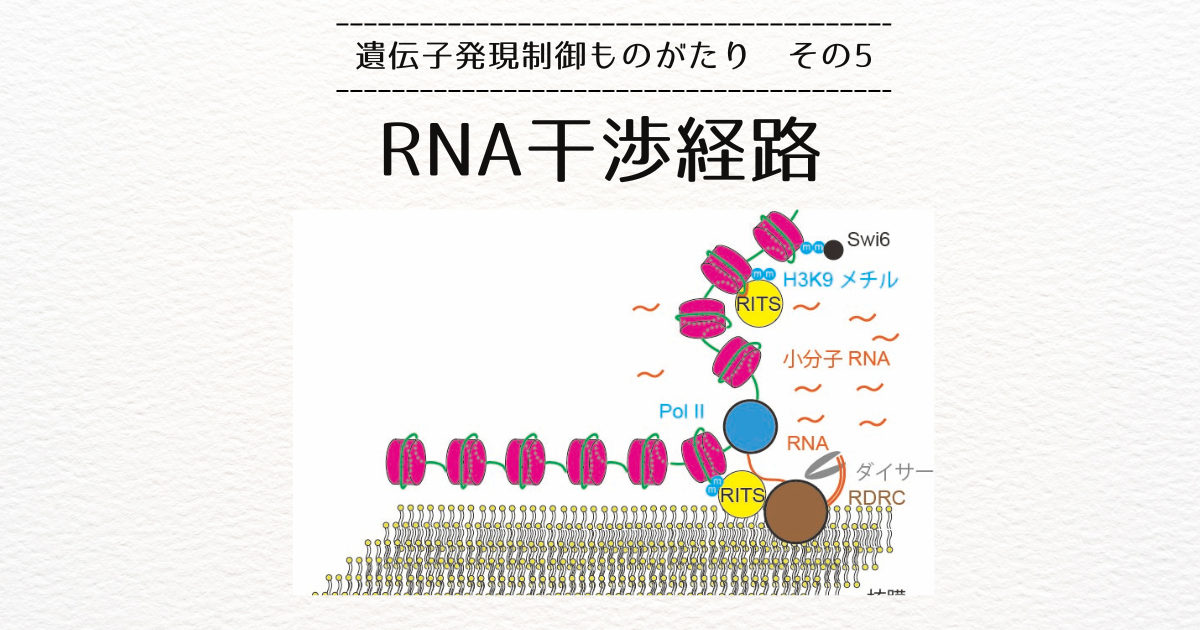
トポロジカルドメインの両端には、コヒーシンとCTCFという二種類のタンパク質がついていることが分かりました。
コヒーシンはリング状のタンパク質ですので、それがクロマチンをまとめてループを作っていると考えられます。
CTCFはDNAに結合するタンパク質で、結合する向きがあります。
ドメインの両端についているCTCFは、互いにドメインの内側を向いているという特徴があります。
特殊な部位で形成されるほかの構造もありますが、
ヌクレオソーム < トポロジカルドメイン < コンパートメント < 染色体テリトリー
が染色体の階層構造の概観となります。
トポロジカルドメインの端で見つかったコヒーシンですが、SMC(Structural Maintenance of Chromosome)タンパク質という染色体の構造を形作るタンパク質の1つです。
リング状というのはだいぶデフォルメがかかっていて、もう少し解像度を高くして言うと、布団をほす洗濯ばさみののような形と思ってもらった方がいいかもしれません。
それでもだいぶデフォルメがかかっていますけどね。
しかし、リングのように閉じた構造だけでなく、開いた構造にもなれるというのがポイントです。
コヒーシンやコンデンシンがDNAを内包するためには(topological loading)、ATPが必要で、それが洗濯ばさみの開閉に関わっていると考えられます。
その他にも、コンデンシンやSMC5/6などもSMCタンパク質のなかまです。
コンデンシンもリング状(洗濯ばさみ状)のタンパク質で、二本のDNAを内包することによってループを形成します。
2012年にJohn Markoが、コンデンシンは分子モータで、内包するDNAを一方向に輸送する活性を持つ(ループ押し出し活性)ことを理論的に予言しています[14]。
コンデンシンが分子モータであることは、当時ブラウン大学にいらっしゃった寺川さんがDNAカーテンという系を用いて一分子観察で示しています[15]。
その後、Cees Dekkerらのグループがコンデンシンのループ押し出し活性の一分子観察に成功しています。
これからお話しするのは、コンデンシンのループ押し出し活性が発見される前にコヒーシンの研究で起きたことです。
トポロジカルドメインがループであることが報告された2年後に、そのメカニズムを提案する論文が2本発表されました[17,18]。
コヒーシンがループ押し出し活性を持っていることと、ループ押し出しが内向きのCTCFに留められることを仮定すると、トポロジカルドメインの接触頻度を再現できることがシミュレーションで示されたのです。
Hi-Cの結果から面白いことを見つけたいと思っていましたので、やられたと思いました。
とはいえ、Hi-Cで何を測っているのかこの論文でやっと分かったというレベルでしたので、当時の私では話にもなりませんでした。
ループ押し出しの論文は、MirnyらとLieberman Aidenらのグループがほぼ同時に発表したのですが、すこし考え方の差があります。
Lieberman Aidenはトポロジカルドメインをループの集合体と考えていたようです[18]。
一方、Mirnyらはトポロジカルドメインは、あくまでも多数の細胞におけるクロマチン構造の平均を取ったために見えるものであり、本質的にはクロマチンループがあるに過ぎないと考えていたようで[17]、後にsingle-cell Hi-Cを使ってそれを示しています[19]。
ループ押し出し理論では、コヒーシンが分子モータであるということは、あくまでも仮定です。
3つのグループがDNAにロードされたコンデンシンの運動の一分子観察を行い、コヒーシンのモータ活性を確かめましたが、双方向のブラウン運動しかしないという結論となっています[20-22]。
そこで、私たちは反撃に出ることにしました。
コヒーシンが分子モータでないとしたら、どのようなメカニズムでループ押し出しが起こるのでしょうか?
コヒーシンはリング状のタンパク質であり、DNAを内包することによってループを作ることは、前述のとおりです。
合成高分子の世界に、スライドリングゲルというものがあります。
スライドリングゲルは、リング状の分子をたくさん持っている高分子(ポリロタキサン)を使って、その一部のリングを二量体化することによって作られるゲルです。
その名の通り、リング状の分子は高分子をスライドすることができます。
二量体は高分子を架橋してゲル化する役割があるのに対して、単量体は高分子上を熱運動して浸透圧を発生します。
この浸透圧がスライドリングゲルの弾性の原因となっていると考えられています[23]。
コヒーシンは、ローダータンパク質であるNIPBL/Mau2によってDNAに搭載されます。
当時は、コヒーシン1つでループを作ることができるという考え方もあったと思いますが、コヒーシンのリングは比較的小さく、DNA一本しか通さないので、ループを作るためには二量体を作る必要があるという考え方が主流だったと思います。
私は、コヒーシンがローダーにたまたま2つ同時についた時には二量体としてDNAにロードされ、DNAループを作るのに対して、コヒーシンが1つだけしかつかなかった場合には単量体としてDNAにロードされ、浸透圧を発生すると考えました[24]。
つまり、単量体のコヒーシンの熱運動によって発生する浸透圧をエンジンとして、二量体がループ押し出しを行うと考えたわけです。
驚いたことに、Davide Marenduzzoらのグループが二量体のみを考えたバージョンのシミュレーションをほぼ同時に報告しています[25]。
そこではじめて、Leonid MirnyのグループとDavide Marenduzzoのグループが私の競争相手だということを知りました。
MirnyやMarenduzzoは自分のグループを持っていますし、おそらく、自分の興味に従って自由に研究できる環境を持っているでしょう。
一方、私はグループをもっておらず、少なくとも2016年までは自分の研究ができる時間は早朝と土日だけだったわけで、こんな状況でどうやって彼らに勝てばいいのだろうと、自分の置かれていた状況が心底嫌になったのを覚えています。
ループ押し出しの浸透圧機構は結構自信があったのですが、本質的に同じアイデアを他の人が独立に思いつくことができるのであれば、オリジナリティーは弱いと言わざるを得ません。
そのため、私は浸透圧機構の研究をつづけることなく、代わりに、ループ押し出しによってクロマチンの運動法則がどのようなものになるかということを調べる研究に舵を切ることとなります[26]。
2019年に、Jan-Michael Petersのグループが、cohesinのループ押出しを一分子観察することに成功しました[27]。
Cohesinがループ押し出し運動をするためには、ローダーであるNIPBL/Mau2が必須という結論でした。
結局、浸透圧機構も机上の空論に終わったこととなります。
この浸透圧機構の論文を通したことによって、様々なSMCタンパク質の研究者と出会うことができました。
また、この論文を契機に科研費にも採択されるようになり、また、さきがけのゲノム合成領域にも採択され、私の研究は質的に変わっていくこととなります。
前回のブログと同じ結論ですが、ちゃんと論文を出すことは、人と出会い、成長し、よりよい研究をするのに重要なのだと思います。
アインシュタインは、”A person who never made a mistake never tried anything new.”(間違えを犯したことのない人間は、新しいことに挑戦しない人間だ)という格言を残しています。
名古屋大学は、この格言を心の支えとしていた時代でした。
1.T. Cremer & C. Cremer, “Chromosome territories, nuclear architecture and gene regulation in mammalian cells”, Nat. Rev. Gen., 2: 292–301 (2001). doi: https://doi.org/10.1038/35066075
2.T. Misteli, “Chromosome territories: The arrangement of chromosomes in the nucleus.”, Nature Education 1:167 (2008). Chromosome Territories | Learn Science at Scitable
3.A. Grosberg, Y. Rabin, S. Havlin, & A. Neer, “Crumpled Globule Model of the Three-Dimensional Structure of DNA”, Europhys. Lett., 23: 373 (1993). doi: https://doi.org/10.1209/0295-5075/23/5/012
4.A. Rosa & R. Everaers, “Structure and Dynamics of Interphase Chromosomes”, PLoS Comput. Biol., 22: e1000153 (2008). doi: https://doi.org/10.1371/journal.pcbi.1000153
5.C. Hoencamp et al., “3D genomics across the tree of life reveals condensin II as a determinant of architecture type”, Science, 372: 984-989 (2021). doi: https://doi.org/10.1126/science.abe2218
6.E. Lieberman-Aiden, N. L. van Berkum, L. Williams, M. Imakaev, T. Ragoczy, A. Telling, I. Amit, B. R. Lajoie, P. J. Sabo, M. O. Dorschner, R. Sandstrom, B. Bernstein, M. A. Bender, M. Groudine, A. Gnirke, J. Stamatoyannopoulos, L. A. Mirny, E. S. Lander, & J. Dekker, “Comprehensive mapping of long-range interactions reveals folding principles of the human genome”, Science, 326: 289-93 (2009). doi: https://doi.org/10.1126/science.1181369
7.T. H. S. Hsieh, ∙ C. Cattoglio, ∙ E. Slobodyanyuk, ∙ A. S. Hansen, ∙ O. J. Rand,∙ R. Tjian, & ∙ X. Darzacq, “Resolving the 3D Landscape of Transcription-Linked Mammalian Chromatin Folding”, Mol. Cell, 78: 539-553 (2020). doi: https://doi.org/10.1016/j.molcel.2020.03.002
8.N. Krietenstein, ∙ S. Abraham,∙ S. V. Venev, ∙ N. Abdennur, ∙ J. Gibcus, ∙ T. H. S. Hsieh, ∙ K. M. Parsi, ∙ L. Yang, ∙ R. Maehr, ∙ L. A. Mirny, ∙ J. Dekker, & ∙ O. J. Rando, “Ultrastructural Details of Mammalian Chromosome Architecture”, Mol. Cell, 78: 554-565 (2020). doi: https://doi.org/10.1016/j.molcel.2020.03.003
9.T. J. Stevens, D. Lando, S. Basu, L. P. Atkinson, Y. Cao, S. F. Lee, M. Leeb, K. J. Wohlfahrt, W. Boucher, A. O’Shaughnessy-Kirwan, J. Cramard, A. J. Faure, M. Ralser, E. Blanco, L. Morey, M. Sansó, M. G. S. Palayret, B. Lehner, L. Di Croce, A. Wutz, B. Hendrich, D. Klenerman & E. D. Laue, “3D structures of individual mammalian genomes studied by single-cell Hi-C”, Nature, 544: 59–64 (2017). doi: https://doi.org/10.1038/nature21429
10.M. Zheng, S. Z. Tian, D. Capurso, M. Kim, R. Maurya, B. Lee, E. Piecuch, L. Gong, J. J. Zhu, Z. Li, C. H. Wong, C. Y. Ngan, P. Wang, X. Ruan, C. L. Wei & Y. Ruan, “Multiplex chromatin interactions with single-molecule precision”, Nature, 566: 558–562 (2019). doi: https://doi.org/10.1038/s41586-019-0949-1
11.J. R. Dixon, S. Selvaraj, F. Yue, A. Kim, Y. Li, Y. Shen, M. Hu, J. S. Liu & B. Ren, “Topological domains in mammalian genomes identified by analysis of chromatin interactions”, Nature, 485: 376–380 (2012). doi: https://doi.org/10.1038/nature11082
12.E. P. Nora, B. R. Lajoie, E. G. Schulz, L. Giorgetti, I. Okamoto, N. Servant, T. Piolot, N. L. van Berkum, J. Meisig, J. Sedat, J. Gribnau, E. Barillot, N. Blüthgen, J. Dekker & E. Heard, “Spatial partitioning of the regulatory landscape of the X-inactivation centre”, Nature, 485: 381–385 (2012). doi: https://doi.org/10.1038/nature11049
13.S. S P Rao, M. H. Huntley, N. C Durand, E. K. Stamenova, I. D. Bochkov, J. T. Robinson, A. L. Sanborn, I. Machol, A. D. Omer, E. S. Lander, E. Lieberman Aiden, “A 3D map of the human genome at kilobase resolution reveals principles of chromatin looping”, Cell, 159: 1665-80 (2014). doi: https://doi.org/10.1016/j.cell.2014.11.021
14.E. Alipour & J. F. Marko, “Self-organization of domain structures by DNA-loop-extruding enzymes”, Nucleic Acids Res., 40: 11202–11212 (2012). doi: https://doi.org/10.1093/nar/gks925
15.T. Terakawa, S. Bisht, J. M. Eeftens, C. Dekker, C. H. Haering, & E. C. Greene, “The condensin complex is a mechanochemical motor that translocates along DNA”, Science, 358: 672-676 (2017). doi: https://doi.org/10.1126/science.aan6516
16.M. Ganji, I. A. Shaltiel, S. Bisht, E. Kim, A. Kalichava, C. H. Haering, and C. Dekker, “Real-time imaging of DNA loop extrusion by condensin”, Science, 360: 102-105 (2018). doi: https://doi.org/10.1126/science.aar7831
17.G. Fudenberg, M. Imakaev, C. Lu, A. Goloborodko, N. Abdennur, L. A. Mirny, “Formation of Chromosomal Domains by Loop Extrusion”, Cell Rep. 15: 2038-2049 (2016). doi: https://doi.org/10.1016/j.celrep.2016.04.085
18.A. L. Sanborn, S. S. P. Rao, S. C. Huang, N. C. Durand, M. H. Huntley, A. I. Jewett, I. D. Bochkov, D. Chinnappan, A. Cutkosky, J. Li, K. P. Geeting, A. Gnirke, A. Melnikov, D. McKenna, E. K. Stamenova, E. S. Lander, and E. Lieberman Aiden, “Chromatin extrusion explains key features of loop and domain formation in wild-type and engineered genomes”, Proc. Natl. Acad. Sci. USA, 112: E6456-E6465 (2015). doi: https://doi.org/10.1073/pnas.1518552112
19.W. Schwarzer, N. Abdennur, A. Goloborodko, A. Pekowska, G. Fudenberg, Y. Loe-Mie, N. A. Fonseca, W. Huber, C. H. Haering, L. Mirny & F. Spitz, “Two independent modes of chromatin organization revealed by cohesin removal”, Nature, 551: 51–56 (2017). doi: https://doi.org/10.1038/nature24281
20.J. Stigler, G. Ö. Çamdere, D. E. Koshland, & E. C. Greene, “Single-Molecule Imaging Reveals a Collapsed Conformational State for DNA-Bound Cohesin”, Cell Rep., 15: 988-998 (2016). doi: https://doi.org/10.1016/j.celrep.2016.04.003
21.I. F. Davidson, D. Goetz, M. P. Zaczek, M. I. Molodtsov, P. J. Huis in ‘t Veld, F. Weissmann, G. Litos, D. A. Cisneros, M. O. Hafalla, R. Ladurner, F. Uhlmann, A. Vaziri & J. M. Peters, “Rapid movement and transcriptional re‐localization of human cohesin on DNA”, EMBO J., 35: 2671–2685 (2016). doi: https://doi.org/10.15252/embj.201695402
22.M. Kanke, E. Tahara, P. J Huis in’t Veld & T. Nishiyama, “Cohesin acetylation and Wapl-Pds5 oppositely regulate translocation of cohesin along DNA”, EMBO J., 35: 2686–2698 (2016). doi: https://doi.org/10.15252/embj.201695756
23.K. Mayumi, M. Tezuka, A. Bando, & K. Ito, “Mechanics of slide-ring gels: novel entropic elasticity of a topological network formed by ring and string”, Soft Matter, 8: 8179-8183 (2012). doi: https://doi.org/10.1039/C2SM25508A
24.T. Yamamoto & H. Schiessel, “Osmotic mechanism of the loop extrusion process”, Phys. Rev. E, 96: 030402(R) (2017). doi: https://doi.org/10.1103/PhysRevE.96.030402
25.C. A. Brackley, J. Johnson, D. Michieletto, A. N. Morozov, M. Nicodemi, P. R. Cook, and D. Marenduzzo, “Nonequilibrium Chromosome Looping via Molecular Slip Links”, Phys. Rev. Lett., 119: 138101 (2017). doi: https://doi.org/10.1103/PhysRevLett.119.138101
26.T. Yamamoto, T. Sakaue, & H. Schiessel, “Loop extrusion drives very different dynamics for Rouse chains in bulk solutions and at interfaces”, Europhys. Lett., 127, 38002 (2019). doi: https://doi.org/10.1209/0295-5075/127/38002
27.I. F. Davidson, B. Bauer, D. Goetz, W. Tang, G. Wutz, and J. M. Peters, “DNA loop extrusion by human cohesin”, Science, 366: 1338-1345 (2019). doi: https://doi.org/10.1126/science.aaz3418