クロマチンの相分離と細胞分化
tetsu
遺伝子発現制御理論物理学研究室
まず、私が遺伝子発現制御の問題を理論物理学でアプローチする研究をするきっかけとなったのは、イスラエルのヴァイツマン研究所での研究経験です。
ヴァイツマン研究所では、所内の実験の研究者ならばだれとでもいいから、共同研究をしてなにか成果を上げなさいという、比較的自由度の高いミッションが与えられました。
その前にいたドイツのMax Planck研究所でも同様なミッションだったので、海外の国立研究所で研究員(ポスドク)をすると、わりと自由度が高いのだと思います。
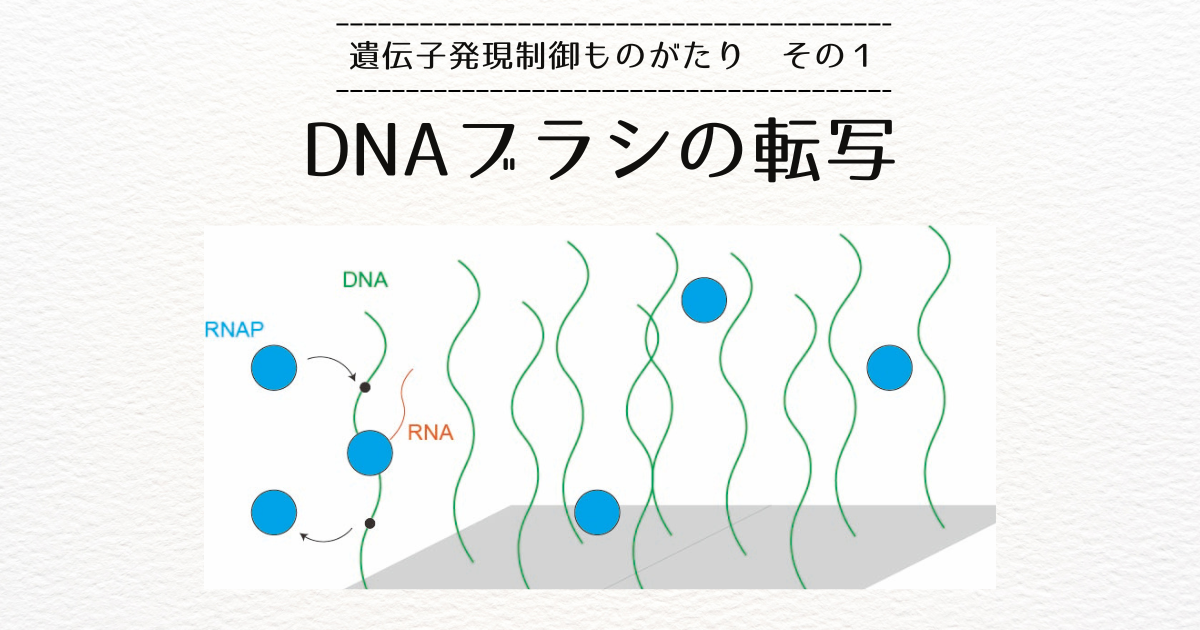
この研究所には、Roy Bar-Zivさんという先生がいらっしゃいまして、たくさんのDNAの一方の先端を表面につけてつくられる「DNAブラシ」という系を作成し、RNA合成の反応速度(転写レート)を測定されていました[1]。
このように生体システムを作って調べる研究を合成生物学といいます。
DNAブラシは生物のどの状況に対応するのかすらわからないような系ですが、遺伝子発現に対する非常に重要な示唆を与えてくれました。
結論から言ってしまうと、DNAの構造が転写レートを制御すると同時に、転写自体が系の構造を制御するということです。
私は、この問題に対し、ソフトマター物理学と化学反応速度論を組み合わせるというアプローチをとりました。
分子生物学者たちが実験的に明らかにしてきたことを見てみると、遺伝子発現制御にDNAの構造や運動がかかわるケースが結構多いことに気付きます。
これがソフトマター物理学を使って遺伝子発現制御の問題に取り組もうと思ったきっかけです。
Bar-Ziv先生が作成したDNAブラシは、ウィルスやバクテリア由来のDNAとRNAポリメラーゼ(RNAP)が使われています。
バクテリアだと制御因子が少なくて済みますので、シンプルな設計が可能なのです。
少し復習すると、転写はRNAPが転写開始配列(プロモータ)に結合し、転写終結配列(ターミネータ)の方向に動きながらDNA塩基と相補的なRNAを合成する過程でした。
プロモータとターミネータの間の部分を転写ユニットと言います。
遺伝子編集によってさまざまなDNAを用意することができるのですが、彼は転写ユニットを1つだけもつDNAを使いました。
転写ユニットとなっているのはDNAの一部なのですが、他の部分は転写とは関係のないただの高分子としてふるまうと考えていいと思います。
RNAPはプロモータからターミネータの方向に動きますが、その方向を転写ユニットの「向き」と定義することができます。
遺伝子編集によって、転写ユニットの(DNA中の)位置と(DNAをつける表面に対する)向きを制御することができます。
また、表面につけるDNAの数(表面積)を変えて、DNAの密度(混雑度)を制御することもできます。
まず、DNAの混雑度が転写ダイナミクスに与える影響を調べるために、DNAの表面密度を大きくしながら転写レートを測定しました。
DNAの表面密度がまだ小さい時には、DNAの表面密度に比例して転写レートが大きくなります。
もちろん、DNAの数が増えるので。
表面密度が大きくなると、だんだん比例からずれてきて、転写レートが飽和することが分かりました。
次に、転写ユニットの位置を、DNAの自由端(表面についていない方の端)、中央、固定端(表面についている方の端)にした場合を比べると、ほとんど転写レートが変わらないことが分かりました。
さらに、転写ユニットが自由端を向いている場合と固定端を向いている場合を比べると、自由端を向いている場合と比べて固定端を向いている方が2倍程度転写レートが大きくなることが分かりました。
DNAブラシの元ネタは、高分子の片方の端を表面につけて作成される高分子ブラシです。
高分子は繰り返し単位であるモノマーをたくさん繋げた非常に長い分子です。
しかし、溶液中では熱運動によってくしゃくしゃになり、割と小さい球形の領域に収まっています。
定量的な説明は他の機会でさせていただきます。
ブラシでは、先端が無理やり表面につけられているので、高分子がある程度伸びた状態になっています。
最初に理論的に示したのがShlomo Alexanderです[2]。
Alexanderはブラシ内部のモノマー濃度が一様であると仮定して計算しました。
このアプローチは、正しい結果を与えるのですが、2つのエラーがたまたまキャンセルするという少し心配なものでもありました。
一方、de Gennesは、自らが作った高分子溶液のスケーリング理論を高分子ブラシに適用し、同じ結論を得ました[3]。
そのため、この理論はAlexander理論、または、Alexander-de Gennes理論と呼ばれます。
高分子ブラシの本質を理解したい人は、de Gennesの論文[3]を読むことをお勧めします。
少し専門的になってしまいますが、表面における高分子の結合点間の距離が、高分子溶液を特徴づける量である相関長となるというのがde Gennesの見つけたことです。
その後、Scott Milner, Tom Witten, Michael Catesによって、平均場理論という方法でブラシ内の濃度プロファイルが表面からの距離の二次関数となっていることが示されます[4]。
ロシアのZurinaさんたちもほぼ同時にこの結論に至ったのですが、参考文献が見つからないので、また後で補完します。
DNAブラシの転写ダイナミクスを調べるために、私はRNAPの濃度分布を解析することにしました[5]。
今言っているRNAPの濃度分布は、ブラシの中の高分子の間の溶液中で熱拡散しているRNAPの濃度分布です。
RNAPの濃度分布は、熱拡散とDNAとRNAPの間の相互作用だけでなく、転写にも影響を受けると考えられます。
熱拡散とDNA-RNAP相互作用だけだったら、Smoluchowski(スモルコウスキー)方程式という良く知られた方程式で定式化することができます。
転写はどのように組み込めばよいでしょうか。
転写の過程では、RNAPがプロモータに結合し、そのRNAPが一定時間後、同じDNAのターミネータから放出されます。
この過程を、RNAPが一定のレートでプロモータの位置でなくなり(シンク)、同じレートでターミネータの位置で発生する(ソース)と読み替えます。
ソースは水の蛇口、シンクはそのまま流しのことです。
出した水がそのまま違う場所で流されていくイメージですね。
私は、このシンクとソースのペアを、「転写ダイポール」と名付けました。
なんでこんなことをしたかというと、実は、誘電分極という電気がたまる現象と転写に共通する本質があることに気付いたからです。
誘電分極は、コンデンサやメモリの原理で、物質(絶縁体)に電圧を加えると電気がたまる現象です。
すると、誘電分極の法則を応用すると、転写によるRNAPの移動をカウントすることができて、それをSmoluchowski方程式に組み込むことができます。
この方法を使って、DNAブラシの転写ダイナミクスの解析を行いました。
DNAの表面密度が大きくなると、ブラシ内部のDNA濃度が高くなります。
その結果、RNAPとの体積排除相互作用によってブラシ内部のRNAPの濃度が小さくなり、転写効率が悪くなることが分かりました。
これがDNAの表面密度を大きくすると転写レートが飽和する原因です。
また、転写ユニットの向きが表面側を向いていると、転写によってRNAPが表面の方に運ばれるので、表面付近のRNAP濃度が高くなります。
逆に、転写ユニットの向きが反対向きだと、転写によってRNAPが外側に運び出されてしまうので、表面付近のRNAP濃度が低くなってしまいます。
そのために、転写ユニットの向きが表面側を向いている方が、反対向きよりも転写レートが大きくなるのです。
この理論を見て、当時の私のボスのSamuel Safran先生に、お前の理論には化学反応が入っているので嫌いだと言われてしまいました。
今考えると、ボスにお前の理論は嫌いだと言われたので、へこむのが普通かもしれませんが、当時の私は、物理の人が化学反応が苦手なら、ソフトマター物理と化学反応を組み合わせる課題は私がやろうと思いました。
また、誘電物理が使えると分かっている系に誘電物理を使って問題を解くのであれば、だれでもできます。
しかし、誘電物理を使って転写の問題を解いたのは、たぶん私がはじめてでしょう。
他の人が誘電分極とは関係ないと思っている問題から誘電分極の本質を見つけ出すことこそが誘電分極のエキスパートの仕事だということも学びました。
DNAブラシは、実際の細胞の状況と非常に異なると思われたかもしれません。
しかし、その理論を作ることで得た、遺伝子発現の頻度は遺伝子の周りの構造によって制御される、遺伝子発現自体が系の構造に影響を与える、という知見は、実際の細胞での遺伝子発現の研究をするうえで重要な視点になると考えました。
その視点で、分子生物学者が明らかにしてきた遺伝子発現制御のメカニズムを見てみると、細胞内の様々な構造が遺伝子発現を制御する例が豊富にあることに気づきます。
ソフトマター物理と化学反応速度論を組み合わせるというアプローチは、実際の細胞の遺伝子発現制御のメカニズムを調べる上でも有用ということです。
幸いにも、ソフトマターの自己組織化(放っておいても勝手に分子が集まって構造が作られる現象)をヴァイツマン研究所で、その後、非平衡ダイナミクス(運動をかっこよく言うとこうなる)も習得することができましたので、遺伝子発現制御を理論物理のアプローチで解き明かすための基礎スキルも身に着けることができました。
名古屋大で助教に採用されたとき、比較的自由に研究させていただける環境だったので、遺伝子発現制御の問題にチャレンジしてみたというわけです。
1.S. S. Daube, D. Bracha, A. Buxboim, & R. H. Bar-Ziv, “Compartmentalization by directional gene expression”, Proc. Natl. Acad. Sci. USA, 107: 2836-2841 (2010). doi: https://doi.org/10.1073/pnas.0908919107
2.S. Alexander, “Adsorption of chain molecules with a polar head a scaling description”, J. Phys. France, 38: 983 – 987 (1977). doi: https://doi.org/10.1051/jphys:01977003808098300
3.P. G. de Gennes, “Conformations of Polymers Attached to an Interface”, Macromolecules, 13: 1069–1075 (1980). doi: https://doi.org/10.1021/ma60077a009
4.S. T. Milner, T. A. Witten, & M. E. Cates, “Theory of the grafted polymer brush”, Macromolecules, 21: 2610-2619 (1988). doi: https://doi.org/10.1021/ma00186a051
5. T. Yamamoto & S. A. Safran, “Transcription rates in DNA brushes”, Soft Matter, 11: 3017-3021 (2015). doi: https://doi.org/10.1039/C4SM02871F