RNA干渉経路
tetsu
遺伝子発現制御理論物理学研究室
ヒトやマウス、植物などの細胞や酵母などのように、細胞内に核を持つ生物を真核生物と言います。
ヒトのDNAは全長2メートルということを聞いたことがあるかと思います。
高分子物理によると、溶液中の高分子は、熱運動のためにくしゃくしゃになり、球状の領域を占めることが分かります。
ヒトの染色体が占める領域は、数を40本として(本当は46本)計算して、1本あたり直径70 マイクロメートル、全部で直径0.3ミリメートル程度です。
これほどの大きさのDNAは、核のようなマイクロメートルの空間にどのように収まっているのでしょうか。
真核生物のDNAは、ヒストンというタンパク質と複合体を形成しています。
この複合体をクロマチンと言います。
ここで言う複合体とは、ヒストンがDNAについている程度に思っていただければ大丈夫です。
後ほど詳しい構造を説明させていただきます。
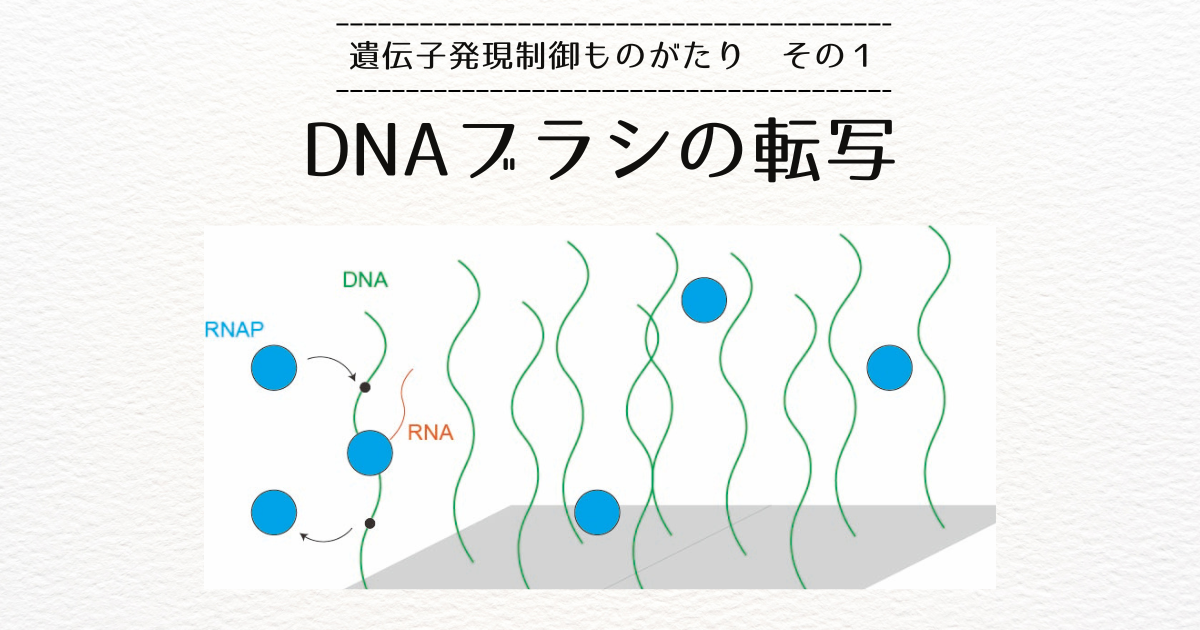
前回お話ししたDNAブラシでは、ヌクレオソームを形成していない裸のDNAを扱っていました[1]。
私はこのブラシの理論を真核生物の転写制御の理解につなげたいと考え、名古屋大学の助教に着任してからは、DNAブラシの理論をクロマチンに拡張することから研究を始めました。
クロマチンを電子顕微鏡で見ると、数珠状の構造が観察されます。
ヒストンには、H2A, H2B, H3, H4の4種類があります。
それが2つずつ合わさって、8量体を形成します。
その8量体にDNAが巻き付いた構造をヌクレオソームと言います。
数珠構造の1つの珠に対応する構造がヌクレオソームです。
それぞれのヒストンには、しっぽ(ヒストンテイル)があります。
ヒストンテイルは、メチル化やアセチル化などの化学修飾を受けます。
化学修飾は、今回のテーマの非常に重要な要素で、細かい分類があるのですが、まずはメチル化やアセチル化されるということだけ知っておいてください。
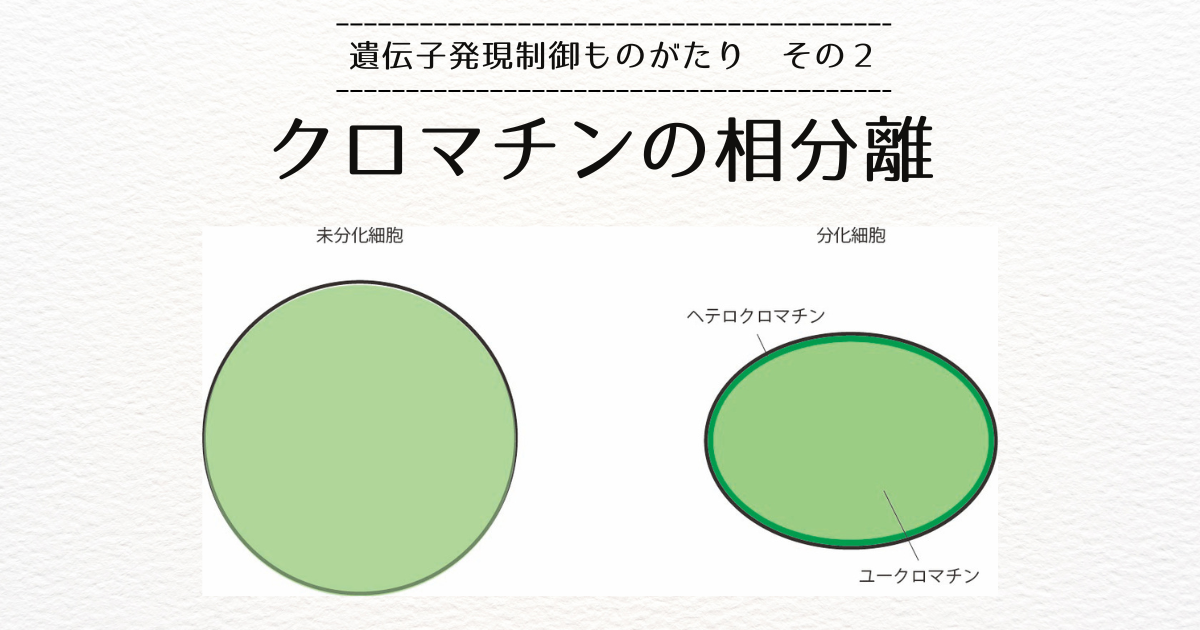
真核生物のDNAには、転写が活性な領域であるユークロマチンと不活性な領域であるヘテロクロマチンがあります。
分化した細胞の多くは、ユークロマチンが核の中央に位置し、ヘテロクロマチンは核膜付近か核小体の周りに観察されます[2]。
ユークロマチンのヌクレオソームでは、ヒストンテイルはアセチル化(H3K27ac)されている傾向があり、ヘテロクロマチンのヌクレオソームでは、ヒストンテイルがメチル化(H3K9me、H3K27me)されている傾向があります。
ここで、アセチル化とかメチル化というのはかなりラフな言い方で、もっと正確にはカッコの中のH3K27acみたいな言い方をしないといけないのです。
たとえば、H3K4meはヒストンテイルのメチル化を表しますが、ユークロマチンで見られる修飾です。
しかし、細かいことを言い始めると難しくなるので、H3K9meとK3K27meをメチル化の代表、H3K27acをアセチル化の代表にさせていただきます。
ちなみに、H3K9meとは、ヒストンH3の9番目のリシン(K)がメチル化されているという意味です。
メチルの数によって、H3K9me、H3K9me2、H3K9me3があります。
同様に、H3K27acは、ヒストンH3の27番目のリシンがアセチル化されているという意味です。
長くなりましたが、重要なのは、ヒストンテイルの修飾と転写の活性状態が紐づいていて、なおかつ、活性状態の同じものは核内の同じ区画に局在化していることです。
また、ユークロマチンのヌクレオソームでは、ヒストンテイルがアセチル化される傾向にあると言いましたが、全体がアセチル化されているというよりは、ごくごくたまにあるというイメージです。
このように、系が二つの領域に分かれる現象を相分離といいます。
たとえば、サラダドレッシングのように、水と油は混ざらずに、水を主成分とする部分と油を主成分とする部分に分かれてしまいますよね。
物理ではそれと同じ現象だととらえるわけです。
前に、多くの分化細胞では、ユークロマチンが核の中央部に位置し、ヘテロクロマチンは核膜付近か核小体付近で観察されるということを言いました。
Irina Soloveiさんたちは、夜行性の動物の目の細胞では、ヘテロクロマチンが中心に局在化していて、ユークロマチンが核膜付近に局在化することを発見しました。
さらに、後続論文では、核膜にあるラミンBレセプタとラミンA/Cをなくすと、夜行性動物の目の細胞以外の分化細胞でもユークロマチンが核膜に位置し、ヘテロクロマチンが中央側に配置されることを報告しています。
この実験で、クロマチンがどうしてこのようなパターンを形成するかということに興味が集まりました。
今となっては、クロマチンのパターンを説明するためのシミュレーションはたくさんあります。
しかし、この問題に最初にシミュレーションのアプローチで答えたのは、私の知る限りMarenduzzoとCookの論文です[5]。
高分子は、同じ繰り返し単位(モノマー)がつながった分子です。
彼らのシミュレーションでは、クロマチンをユークロマチン的な性質を持つモノマーからなる鎖(ブロック)とヘテロクロマチン的な性質を持つモノマーからなる鎖(ブロック)が結合したものだと考えます。
このような異種の高分子をつなげて作られる高分子をブロック共重合体と言います。
彼らは特に転写などというものは考えなくても、熱運動だけで観察されたクロマチンのパターンが形成されることを示しています。
ユークロマチン的、あるいは、ヘテロクロマチン的な性質としては、主に大きさや硬さが考慮に入れられていて、立体排除以外の相互作用は考えていません。
このように、熱運動だけで見かけの引力相互作用が生じる現象を枯渇効果(depletion effect)と言います。
分化した後の細胞では、クロマチンが特徴的なパターンを示すことをご紹介しました。
これらの細胞は受精卵が繰り返し分裂する中でできてくる細胞です。
それでは、初期の未分化の細胞のクロマチンはどのようなパターンを形成しているでしょうか。
実は、未分化細胞のクロマチンは、アセチル化している部分が多く、核内で一様に分布しています[6]。
木村暁さんらの論文では、未分化細胞は比較的球形で体積が大きいのですが、分化するにつれて扁平となり体積も小さくなることが示されています[6]。
DNAの量は一定ですので、分化するにつれてDNAの濃度が高くなるということです。
分化していく間に、ヘテロクロマチン領域の核生成が観察されています[6]。
分化の間にそれぞれの遺伝子の活性が変わっていきますが、ヘテロクロマチンが生成することと無関係ではないはずです。
CookとMarenduzzoのシミュレーションをはじめ、多くのシミュレーションではクロマチンがすでにユークロマチン的なブロックとヘテロクロマチン的なブロックからなることを仮定します。
しかし、一様なクロマチンがどのようにユークロマチン的なブロックとヘテロクロマチン的なブロックに分かれ相分離していくかということが分かれば、細胞分化の間の遺伝子発現制御に迫ることができるのではないでしょうか。
私がこの研究を始めた当初、クロマチンの構造はすでに物理のテーマになっていました。
その理論の第一人者は、当時オランダのライデン大学にいた(2026年2月現在ドレスデン工科大にいる)Helmut Schiesselさんという方です。
このブログでお話しする研究プロジェクトを構想したのは、名大に移る1年前の夏で、Schiesselさんと一緒にEUのマリーキュリーファンディングに応募したときです。
私は転写の研究をしてきたが、真核生物ではクロマチンがどのように転写を制御するか知りたく、私のDNAブラシの理論を拡張したいということをお話ししました。
当時中国にいたので、skypeかなんかで話をしたのだと思いますが、転写の間にRNAポリメラーゼがヌクレオソームと衝突すると、ヒストンがDNAから取れてしまうことを試験管実験で示した実験を紹介していただきました[7]。
ここで、試験管実験とは、生きた細胞内ではなく、DNAとRNAポリメラーゼなどの転写に必要なタンパク質を精製し、精製タンパク質の水溶液中で行った再構成実験のことです。
また、ヒストンは正電荷を持っているのですが、負電荷をもつDNAが過剰に巻かれていて、ヌクレオソームコアの電荷は負となっています。
ヒストンテイルは無修飾の時には正電荷を持っているので、他のヌクレオソームコアの負電荷と引き合うので、ヌクレオソームは互いにくっつき合う性質があることを示すご自身のシミュレーションも教えていただきました[8]。
このような効果をテイルブリッジング効果というそうです。
一方、ヒストンテイルがアセチル化すると、正電荷が無くなるため、静電引力が働かなくなります。
ヌクレオソームがくっつき合う性質を持つことと転写の間にRNAポリメラーゼによってヌクレオソームが壊れることを組み合わせると、クロマチンが相分離という現象を示すことに気付きました。
結局、マリーキュリーファンディングには採択されませんでしたが、プロジェクトが面白いので、名古屋大に移ってからSchiesselさんと共同研究することにしました。
私は、RNAポリメラーゼとヌクレオソームの衝突によるヌクレオソームの分解の過程とヌクレオソームが形成される過程を考慮に入れて、DNAブラシの転写ダイナミクスの理論を拡張しました[9]。
ダイナミクスを語るとき、重要なのはレートです。
ヌクレオソームが形成されるレートよりも転写レートの方が大きい時には、ほぼ裸の膨潤した状態(ユークロマチン)が安定となり、逆の場合には、ヌクレオソームが形成されている収縮した状態(ヘテロクロマチン)が安定になります。
ちょうど、これらのレートが同じくらいの時にクロマチンブラシの相分離が起こることが分かりました。
その後、収縮するDNAは表面近くに存在し、その上の部分を膨潤したDNAがカバーするような構造が安定であるという分化細胞で観察されるものと同じような構造が予言されました[10]。
さらに、この理論を高分子ゲルに拡張し、核が負のポアソン比を示すことを示すことができました[11]。
これまでの説明は、全体的なサイエンスの流れを説明したものです。
しかし、私の視点からすると、Schiesselさんに教えてもらったこと(ヌクレオソーム間の相互作用とRNAポリメラーゼとの衝突によるヌクレオソームの分解)を考慮に入れて、とりあえずDNAブラシをクロマチンに拡張したまでです。
ユークロマチンとヘテロクロマチンが分化細胞の核内で相分離していることだとか、未分化細胞ではクロマチンが一様だとか、そういうことは全部後付けです。
これらの実験的な事実は、自分で論文[2,6]を読んで得た情報です。
一方、Solovei実験[3,4]とCookとMarenduzzoのシミュレーション[5]については、理論を作って論文を書くときにSchiesselさんから教えてもらいました。
研究を始めたときには前提知識をほとんど持っていなかったのに、ある程度つじつまがあう理論ができたのは、単純にミラクルです。
とはいえ、NCCでほとんど時間がとられている中、ほぼ土日だけで一年で1つの仕事を仕上げたので、good enoughだと思いたいです。
作った理論が本当に正しかということに言及していきます。
実は、現在では、東大の胡桃坂さんが、FACTというタンパク質(シャペロン)によって、ヒストンはDNAから完全にはがれるわけではなく、後方に新しいヌクレオソームを作ることをクライオ電子顕微鏡で示しています[12]。
理論を作るのに参考にした試験管実験では、FACTが足りてなかったのです。。。
試験管実験は系が比較的単純なのが長所ですが、必要な因子が足りていない場合があるところが怖いですね。
京大の寺川さんは、むしろRNAポリメラーゼはヌクレオソームを前に押すことを示しています[13]。
RNAポリメラーゼとヌクレオソームの相互作用はそんなに単純ではないようです。
また、ユークロマチン領域は裸のDNAが膨潤しているのではなく、クロマチンを作ったまま膨潤しています。
今では、我々が考案した転写によって駆動されるクロマチン相分離の理論は、机上の空論だったと思っています。
しかし、どこかで使える要素は残っていると思っています。
アセチル化とメチル化の酵素反応ダイナミクスを考慮に入れてクロマチンの相分離を予言する理論を2020年に作りましたが、こちらは雑誌への採択すらされませんでした[14]。
査読者のコメントはあまり納得のいくものではありませんでしたが、結局諦めてしまいました。
この研究を行い、論文を書いたことは、Helmut Schiesselさんだけでなく、新学術木村領域(クロマチン潜在能)に参加し、日本の様々な分子生物学者と出会い、交流し、共同研究を行っていくきっかけになりました。
新しい人と出会うと、成長するきっかけとなります。
一般的には、自分が成長し続けている限り、現在行っている研究よりも、未来に行う研究の方が良いものになる可能性は高くなります。
論文を書くことはそのためのきっかけになりますので、例え希望のジャーナルに掲載される自信がなくてもちゃんと論文にすることは重要なのだと思います。
1.T. Yamamoto & S. A. Safran, “Transcription rates in DNA brushes”, Soft Matter, 11: 3017-3021 (2015). doi: https://doi.org/10.1039/C4SM02871F
2. A. Akhtar & S. Gasser, “The nuclear envelope and transcriptional control”, Nat. Rev. Genet., 8: 507–517 (2007). doi: https://doi.org/10.1038/nrg2122
3.I. Solovei, ∙ M. Kreysing, ∙ C. Lanctôt, ∙ S. Kösem, ∙ L. Peichl, ∙ T. Cremer1, ∙ J. Guck, & B. Joffe, “Nuclear Architecture of Rod Photoreceptor Cells Adapts to Vision in Mammalian Evolution”, Cell, 137: 356-368 (2009). doi: https://doi.org/10.1016/j.cell.2009.01.052
4.I. Solovei, ∙ A. S. Wang, ∙ K. Thanisch, ∙ C. S. Schmidt, ∙ S. Krebs, ∙ M. Zwerger, ∙ T. V. Cohen, ∙ D. Devys, ∙ R. Foisner, ∙ L. Peichl, ∙ H. Herrmann, ∙ H. Blum, ∙ D. Engelkamp, ∙ C. L. Stewart, H. Leonhardt, & ∙ B. Joffe, “LBR and Lamin A/C Sequentially Tether Peripheral Heterochromatin and Inversely Regulate Differentiation”, Cell, 152: 584-598 (2013). doi: https://doi.org/10.1016/j.cell.2013.01.009
5.P. R. Cook & D. Marenduzzo, “Entropic organization of interphase chromosomes”, J. Cell. Biol., 186: 825–834 (2009). doi: https://doi.org/10.1083/jcb.200903083
6.R. Arai, T. Sugawara, Y. Sato, Y. Minakuchi, A. Toyoda, K. Nabeshima, H. Kimura, & A. Kimura, “Reduction in chromosome mobility accompanies nuclear organization during early embryogenesis in Caenorhabditis elegans”, Sci. Rep., 7: 3631 (2017). doi: https://doi.org/10.1038/s41598-017-03483-5
7.L. Bintu, M. Kopaczynska, C. Hodges, L. Lubkowska, M. Kashlev & C. Bustamante, “The elongation rate of RNA polymerase determines the fate of transcribed nucleosomes”, Nat. Struct. Mol. Biol., 18: 1394–1399 (2011). doi: https://doi.org/10.1038/nsmb.2164
8.F. Mühlbacher, C. Holm & H. Schiessel, “Controlled DNA compaction within chromatin: The tail-bridging effect”, Europhys. Lett., 73: 135 (2006). doi: https://doi.org/10.1209/epl/i2005-10351-4
9.T. Yamamoto & H. Schiessel, “Transcription Driven Phase Separation in Chromatin Brush”, Langmuir, 32: 3036–3044 (2016). doi: https://doi.org/10.1021/acs.langmuir.6b00442
10.T. Yamamoto & H. Schiessel, “Transcription dynamics stabilizes nucleus-like layer structure in chromatin brush”, Soft Matter, 13: 5307-5316 (2017). doi: https://doi.org/10.1039/C7SM00239D
11.T. Yamamoto & H. Schiessel, “Chromatin gels are auxetic due to cooperative nucleosome assembly and disassembly dynamics”, Europhys. Lett., 118: 28003 (2017). doi: https://doi.org/10.1209/0295-5075/118/28003
12.H. Ehara, T. Kujirai, M. Shirouzu, H. Kurumizaka, & S. Sekine, “Structural basis of nucleosome disassembly and reassembly by RNAPII elongation complex with FACT”, Science, 377, abp9466. doi: https://doi.org/10.1126/science.abp9466
13.F. Nagae , G. B Brandani , S. Takada, & T. Terakawa, “The lane-switch mechanism for nucleosome repositioning by DNA translocase”, Nucleic Acids Res., 49: 9066–9076 (2021). doi: https://doi.org/10.1093/nar/gkab664
14.T. Yamamoto, T. Sakaue, & H. Schiessel, “Phase separation of chromatin brush driven by enzymatic reaction dynamics of histone posttranslational modifications”, bioRxiv (2020). doi: https://doi.org/10.1101/2020.11.30.405134